Greeting behavior between dogs in a dog park
Abstract
I used naturalistic observations to explore patterns of greeting behavior between 58 off-leash dogs (Canis familiaris) at a dog park. Dogs greeting at a dog park entailed one or both dogs approaching, engaging in olfactory inspection, and then departing or, on occasion, engaging in play. Single-dog greetings, where one dog approached, were statistically more frequent than dual-dog greetings, where both dogs approached. In single-dog greetings, the dog initiating the greeting was significantly bigger and had body posture higher than or equal to that of the recipient dog. Play did not commonly follow greetings, and no incidents of aggression in conjunction with greetings were observed. Voluntary greeting times between dogs in this study were short (mean greet times between unleashed dogs were 6.042 ± 0.801 s). Reciprocated greetings, where both dogs engaged in olfactory inspections of the anogenital region, face/head, and/or inguinal region of the other dog, were not significantly more frequent than unreciprocated greetings, where only one of the dogs engaged in olfactory inspection. However, dogs in reciprocated greetings were more evenly matched in terms of weight than dogs in unreciprocated greetings. In 48.1% of dyads, greeting resulted in a significant change of body posture in one or both dogs. This change was from either a high or low posture at the beginning of the greeting to a neutral posture at the end in most cases. This suggests that greetings in domestic dogs may function in conflict management by reducing arousal and fear. In single-dog greetings, olfactory inspections varied statistically by greeter role (initiator vs. recipient of greeting). The applications of these finding to clinicians working in companion dog behavior are discussed.
Keywords: dog behavior, dog greeting behavior, dog parks, body postures; off-leash dog behavior
Highlights
Dogs greeting one another at a dog park do so as a means of information gathering and conflict management.
Greet times between dogs under semi-naturalistic conditions were relatively short, lasting only a few seconds.
Body postures after a greeting indicated reduced arousal and fear in one or both dogs in approximately half of all greetings.
The findings reported have implications for the management of dogs in public spaces where dog-to-dog encounters may be likely.
Introduction
Researchers have defined greetings in non-human animals as a ritualized pattern of behaviors that occur when animals meet or reunite (Kutsukake et al. 2006; Dias et al. 2008;). Greetings can serve multiple functions including reinforcing group cohesion and social bonds (Smuts and Watanabe 1990; De Marco et al. 2014). For example, female spotted hyenas (Crocuta crocuta) who have been separated will, upon reunion, preferentially direct greetings towards kin and close social companions over less familiar individuals (Smith et al. 2011). Similarly, in a captive pack of 14 African wild dogs (Lycaon pictus), individuals greeted not only when reunited after a separation, but also when together, suggesting that greetings function to promote cooperation among group members (Rütten and Fleissner 2004).
In addition to the promotion of social cohesion and cooperation in some species, greetings also allow individuals to gather information (Howse et al. 2018), assess potential competitors, and manage conflicts without having to resort to fighting (Colmenares et al. 2000; Smuts 2002; Aureli and Schaffner 2007). This is especially important during tense situations in which unfamiliar individuals with the potential to severely injure or kill one another are greeting (Preuschoft and van Schaik 2000). For example, male hamadryas baboons (Papio hamadryas) have large canine teeth used in fighting. Males competing over estrous females use greetings as an assessment strategy to avoid costly aggression (Colmenares 1991). During greetings, animals are in close proximity and vulnerable body parts, such as the penis (in male primates) or the pseudo-penis (in female spotted hyenas) are often exposed and inspected by one or both individuals. When both individuals engage in similar greeting behaviors, the greeting is labeled as “reciprocated”. When one individual ignores, moves away, or responds with aggression, the greeting is labeled as “unreciprocated” (Kummer 1968; Colmenares 1990).
Domestic dogs are also known to engage in greeting behaviors. Dogs who meet and greet will inspect each other’s groin, face, and/or anogenital region (Bradshaw and Lea 1992; Rezac et al. 2011; Howse et al. 2018), areas where pheromones are secreted that allow for intra-specific olfactory communication related to gender, reproduction, and status in dogs and wolves (Harrington and Asa 2003; Pageat and Gaultier 2003). These “inspections” entail some risks because dogs are exposing sensitive body areas, such as their genitals, to the inspecting face and neighboring teeth of other dogs during greetings. Larger individuals may have a size advantage in the advent that a greeting turns tense or leads to aggression, and dogs with higher body postures may convey higher status and be more confident (Hasegawa e al. 2014) during greetings.
Body postures indicate status in dogs (van der Borg et al. 2015) and also serve as a guide to their emotional states (Siniscalchi et al. 2018). For example, high body postures may signal confidence (Petrak 2010), but high postures during a greeting can also indicate over-arousal (Hecht and Horowitz 2015), and unchecked over-arousal in dogs has the potential to tip over into aggression (Trisko and Smuts 2015). In contrast to high postures, low postures in dogs are associated with low status, fear, and anxiety (Schenkel 1967; Wan et al. 2012). With neutral body postures, dogs are at baseline, neither overly fearful nor aroused (Beerda et al. 1998; Kiddie and Collins 2014).
To date, much of the systematic research on greeting behavior has been done in primates and, to a lesser extent, non-domesticated, social carnivores. However, given that there are now more than 76 million pet dogs living in U.S. households (AVMA 2018), and off-leash dog parks are growing faster than any other type of park in America’s largest cities (The Trust for Public Lands 2018), information on canine greeting behaviors in these settings would likely be beneficial. For example, having baseline information on how much time dogs actually spend greeting when they are untethered by leashes and free to move away may be especially relevant to owners looking to manage and/or prevent problematic dog-to-dog encounters.
The objective of the current study was to explore characteristics of dyadic greeting behaviors between dogs meeting at a dog park. I collected naturalistic observations and videotaped greetings between dogs to investigate greeting durations, who initiates (one or both dogs), whether greetings ended in play or aggression, body postures (e.g., neutral, low, or high), and olfactory inspections of the other dog (e.g., anogenital, face and head, or inguinal). I predicted that dogs greeting should act to maximize individual benefits and minimize costs and proposed the following hypotheses: Larger dogs and dogs with higher body postures may be more likely to initiate dyadic greetings and engage in olfactory inspections. Second, olfactory inspections by both dogs (as opposed to one dog) during a greeting should be more common between similarly sized individuals. Third, if greetings function as a form of conflict management in dogs, as they do in other species, body postures after a greeting should indicate a decrease in fear or arousal (i.e., neutral body posture) at the end of the greeting when compared to postures at the beginning of a greeting. To my knowledge, this is the first study to examine body postures as a function of greeting behavior at an off-leash dog park.
methods
Location and Subjects
Observational data on greetings between owned dogs were collected at Swift Run Dog Park in Ann Arbor, Michigan between April 29, 2015 and May 5, 2016. Swift Run is a fenced 10-acre park consisting of a grassy field, a mowed trail, benches, and a single, double-gated entrance into the park.
Data were collected on 58 dogs consisting of 29 unique dyads. Thirty of the dogs were female; the mean age was 3.2 years; 47 were neutered, and 26 were purebred. The weights of the dogs ranged from 2.268 - 43.083 kg with a mean of 25.670 ± 1.303 kg. Dogs were at least 6 months old and with no discernable physical disabilities.
Procedure
Upon entering the park, I positioned myself away from the entrance gate (~ 50 m) and near the periphery of the park boundary to maximize visibility and avoid interfering with the dogs. Greetings were documented using a Sony N 50 video camera fitted with a zoom lens (75-300 mm) and mounted on a tripod. A dyadic greeting occurred when one or both dogs oriented towards and approached by moving within ~ 30 cm of the other, engaged in olfactory inspection, and then departed by turning and moving away (see definitions below).
Dogs were chosen using a non-probability, convenience sampling method (cf. Bauer and Smuts 2007; Horowitz 2009) and videotaped when greetings occurred between any two individuals in the sample population with one exception. Greetings involving dogs just coming through the park entrance gate were not used because these dogs were often converged upon by dogs already at the park (Howse et al. 2018). This commonly caused the new arrivals to act tentatively until they could adjust to the environment. These new arrivals were only considered for inclusion in a greeting after they moved away from the dogs gathered at the gate, and their body posture returned to a more neutral position, indicating that they were not obviously fearful or highly aroused.
Videotaped greetings were later included in the analysis if the interaction was dyadic, the entire greeting was observable, the dogs had not met before (obtained from owner reporting), and human interference was absent.
Owners were queried before or after filming, and all consented to allow their dogs to be included in the study. No identifying owner information was collected, and owners were not recorded. Video recordings were supplemented with owner reports on age, sex, neuter status, breed, weight, and whether their dog had previously met their greeting partner where possible.
Behaviors and Coding
Videos of greetings were downloaded onto a computer and observed in iMovie (Apple Inc.) using real time, slow motion, and frame-by-frame analysis, while the observer coded the time of the behavior (to the nearest s) and/or occurrence of the behavior into an Excel spreadsheet. The following specific information was recorded:
Initiation of Greeting: Either one or both dogs oriented towards and approached to within ~ 30 cm of the other dog. Time noted when dogs were within ~ 30 cm.
Initiator(s): Identity of dog(s) initiating the greeting.
Recipient: Identity of the non-initiating dog in a greeting. The recipient did not approach at the beginning of a greeting.
Termination of Greetings: Time noted when one or both dogs disengaged from the interaction by turning and/or moving away or an initial greeting ended in play.
Duration of Greeting: “Termination of Greeting” - “Initiation of Greeting”
Single-Dog Greetings: Occurred when one (rather than both dogs) initiated a greeting.
Dual-Dog Greetings: Occurred when both dogs (rather than a single dog) initiated a greeting.
Olfactory Inspection: I looked at who initiated olfactory inspections during greetings: the dog initiating the greeting, the recipient, or both dogs. One or both dogs in a greeting would put their nose to the anogenital region (area around the anus and genital region), face/head, and/or inguinal region (groin) of the other dog. This could occur once or multiple times within a greeting. The identity of the dog engaging in olfactory inspection (initiator or recipient) and the location of each inspection were recorded. Multiple inspections were counted when the dog investigated different locations—e.g., moving from face to anogenital region would be counted as two—or when the dog investigated the same region multiple times (with a movement of the nose away from the region in between inspections).
Reciprocated Greetings: Both dogs engaged in olfactory inspection during a greeting.
Unreciprocated Greetings: Only one dog engaged in olfactory inspection during a greeting.
Body Postures: At the initiation and termination of greetings, body postures of both the initiator and recipient (or both initiators in dual-dog greetings) were recorded as “high”, “neutral”, or “low” based on position and carriage of the tails, legs and back, ears, and head (Table 1).
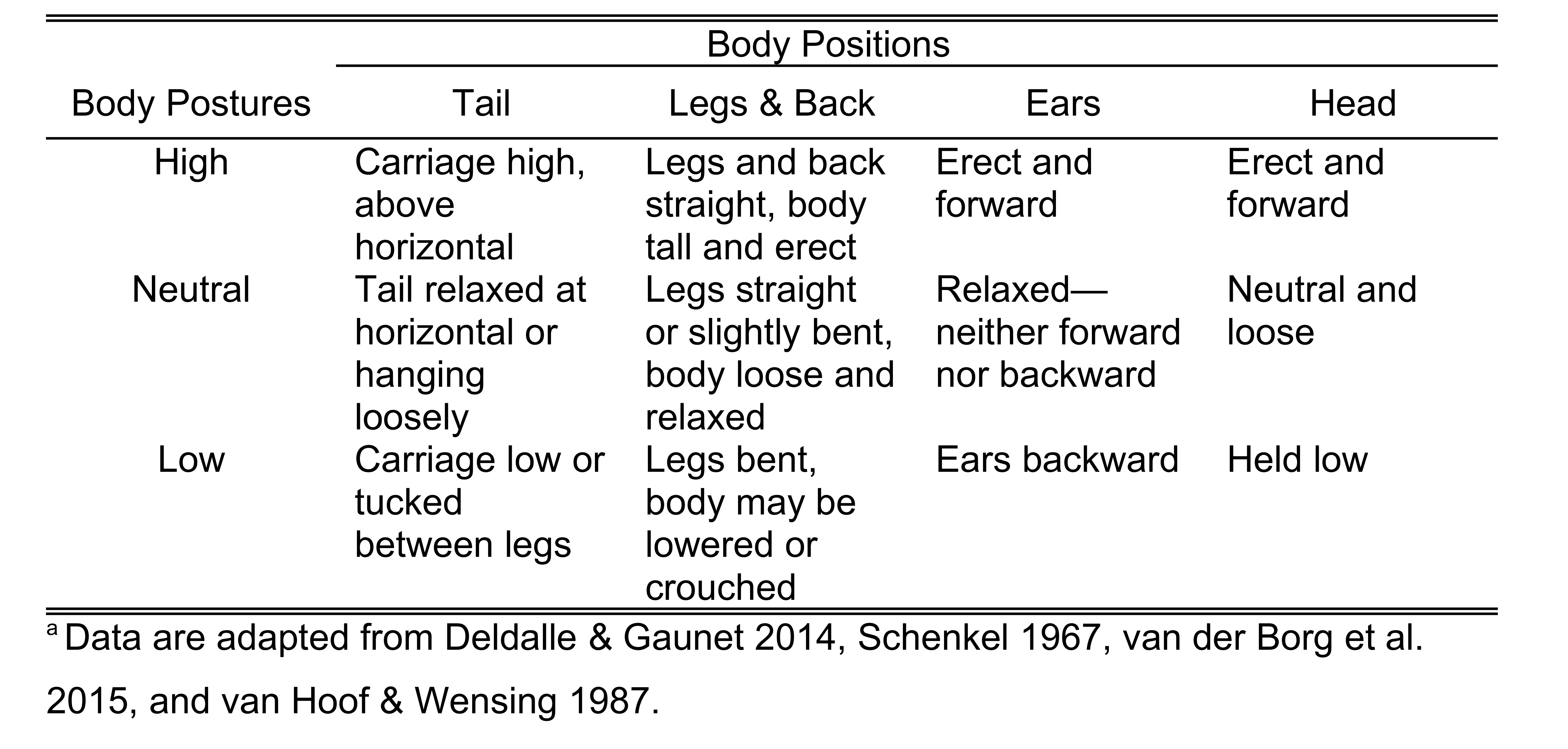
Table 1. Ethogram of body positions of the tail, legs and back, ears, and head as indicators of high, neutral, and low body posturesa
Social Play: Social behavior such as chasing or wrestling often accompanied or was preceded by loose, bouncy movements in one or both dogs (Fagen 1981; Bekoff 1984). Whether or not each greeting dyad played was coded.
Aggressive Behaviors: Threats such as open mouth with teeth displayed accompanied by wrinkling of the face and forehead (van Hooff and Wensing 1987), snaps, bites, fighting (van der Borg et al. 2015). Whether or not dogs engaged in aggressive behaviors was recorded.
Inter-Rater Reliability
The author coded every dyadic greeting from videotapes. A second, trained coder coded a random selection of 20.6% of dyadic greetings to assess inter-rater reliability on 6 variables: 1) initiator, 2) identity of dog(s) engaged in olfactory inspection, 3) location of olfactory inspection (anogenital, face/head, inguinal,), 4 and 5) body postures (low, neutral, high) at the beginning and end of greetings, and 6) length of greeting (coding start and end of each greeting). Percent agreement was 80.0% (Cohen’s Kappa = 0.800) for identity of dog(s) engaged in olfactory inspection (2), location of olfactory inspection (3) and for body postures at the beginning of the greeting (4). Percent agreement was 100% (Cohen’s Kappa = 1.000) for identity of initiator (1) and body postures at the end of the greeting (5). The intraclass correlation coefficient (ICC) for inter-rater reliability using a two-way mixed model with absolute agreement was 0.996 (95% confidence interval: 0.978-0.999) (6). An intraclass coefficient above 0.90 is considered excellent (Perinetti 2018). A Kappa of 0.610-0.800 is substantial, and 0.810-1.000 is almost perfect agreement (McHugh 2012).
Statistical Analysis
Data were analyzed with XLSTAT Software (Addinsoft) using Chi-square test of goodness-of-fit and Binomial tests where noted. When Chi-square tests were significant, Binomial tests were used to test for significance of each group against chance probability (McDonald 2008). This study is largely exploratory in nature. As such, Bonferroni corrections were not used for multiple tests because it results in a reduction of statistical power and an increased likelihood of a Type II error (Nakagawa 2004). A McNemar’s test was run to examine the proportion of dogs in high or low body postures at the beginning of the greeting compared to the body postures of the same dogs at the conclusion of the greeting. Reporting was based on two-tailed tests; alpha was set at 0.05, and no individual was used in the analyses more than once. Small sample size precluded statistical testing by sex and age for this study.
Results
There were no observed incidents of aggression following greetings, and four of the greetings (13.8%) were followed by play. Mean duration of greetings was 6.042 ± 0.801 s.
Single- vs. Dual-Dog Greetings
Single-dog greetings (79.3%) were more common than dual-dog greetings (Binomial test: N = 23 dyads; P = 0.002). In single-dog greetings, the initiator tended to be the heavier of the two dogs (Mann Whitney U Test: U = 146.000; N1 = 23; N2 = 23; P = 0.009; the mean weight for initiators 31.218 ± 1.797 kg vs. 24.020 ± 2.159 kg for recipients) and also displayed body postures that were equal to or higher than that of the recipient dog (Figure 1).
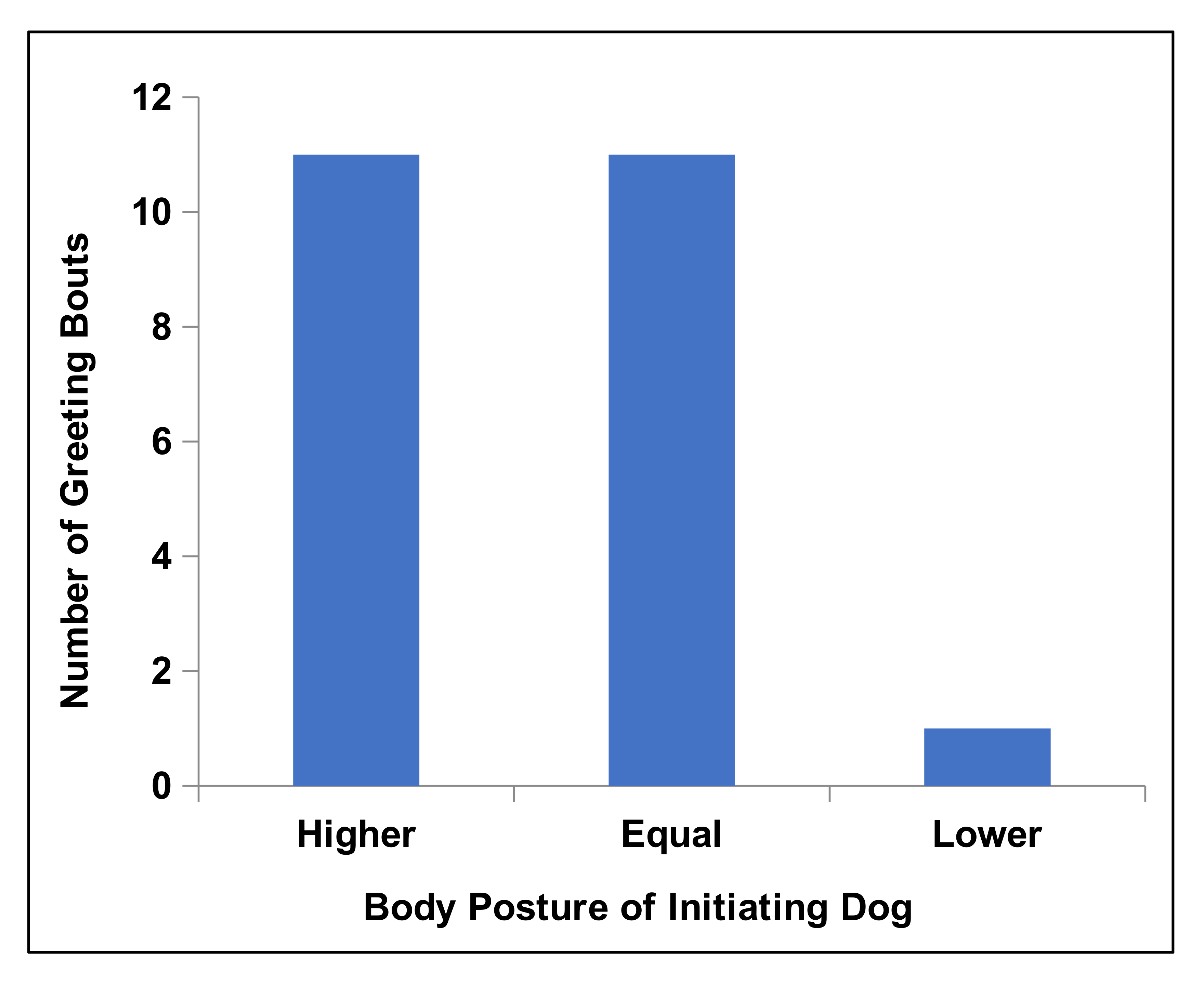
Figure 1. Body posture of dog initiating greeting relative to body posture of recipient dog in single-dog greetings. Measured at the initiation of the greeting. Posture categories are accordingly: high posture > neutral posture > low posture. “Higher” includes all dyads where the dog initiating the greeting was in a higher posture category than the other dog: “Equal” includes all dyads where the two dogs were in the same category; “Lower” includes all dyads in which the dog initiating the greeting was in a lower category than the other dog. Chi-square test of goodness-of-fit:X22 =8.696; P = 0.013.
Reciprocated vs. Unreciprocated Greetings
Sixteen greetings (55.2%) were reciprocated (Binomial test: P = 0.712; no difference in frequency of reciprocated vs. unreciprocated greetings), and participants in reciprocated greetings had smaller mean weight differences — 7.030 ± 1.692 kg — than participants in unreciprocated greetings — 13.668 ± 1.381 kg — (Mann Whitney U Test: U = 165.000; N1 = 16; N2 = 13; P = 0.009).
Body Postures
Sixteen dogs in 13 dyads (48.1% of dyads, body posture data were not available for two dyads) displayed different body postures at the termination of the greetings compared to postures at the initiation of the greeting. In 10 dyads, only one dog (rather than both dogs) shifted posture. Within the 13 dyads where body posture changes occurred, all 11 dogs with high body postures and all four dogs with low body postures at the beginning of the greeting transitioned into neutral postures at the end of the greeting. In one dyad, a dog went from a “neutral” body posture at the beginning of the greeting to a “low” posture at the end. There were no occurrences of a dog moving into a “high” posture at the end of a greeting from another posture.
The proportion of dogs exhibiting neutral body postures at the termination of greetings increased as the proportion of dogs exhibiting either high or low body postures decreased across all dyads for which there was complete body posture data. There was a 47.8% reduction in the number of dogs displaying high body postures, a 56.0% increase in the number of dogs displaying neutral postures, and a 50.0% reduction in the number of dogs displaying “low” body postures at the end of greetings (Figure 2).
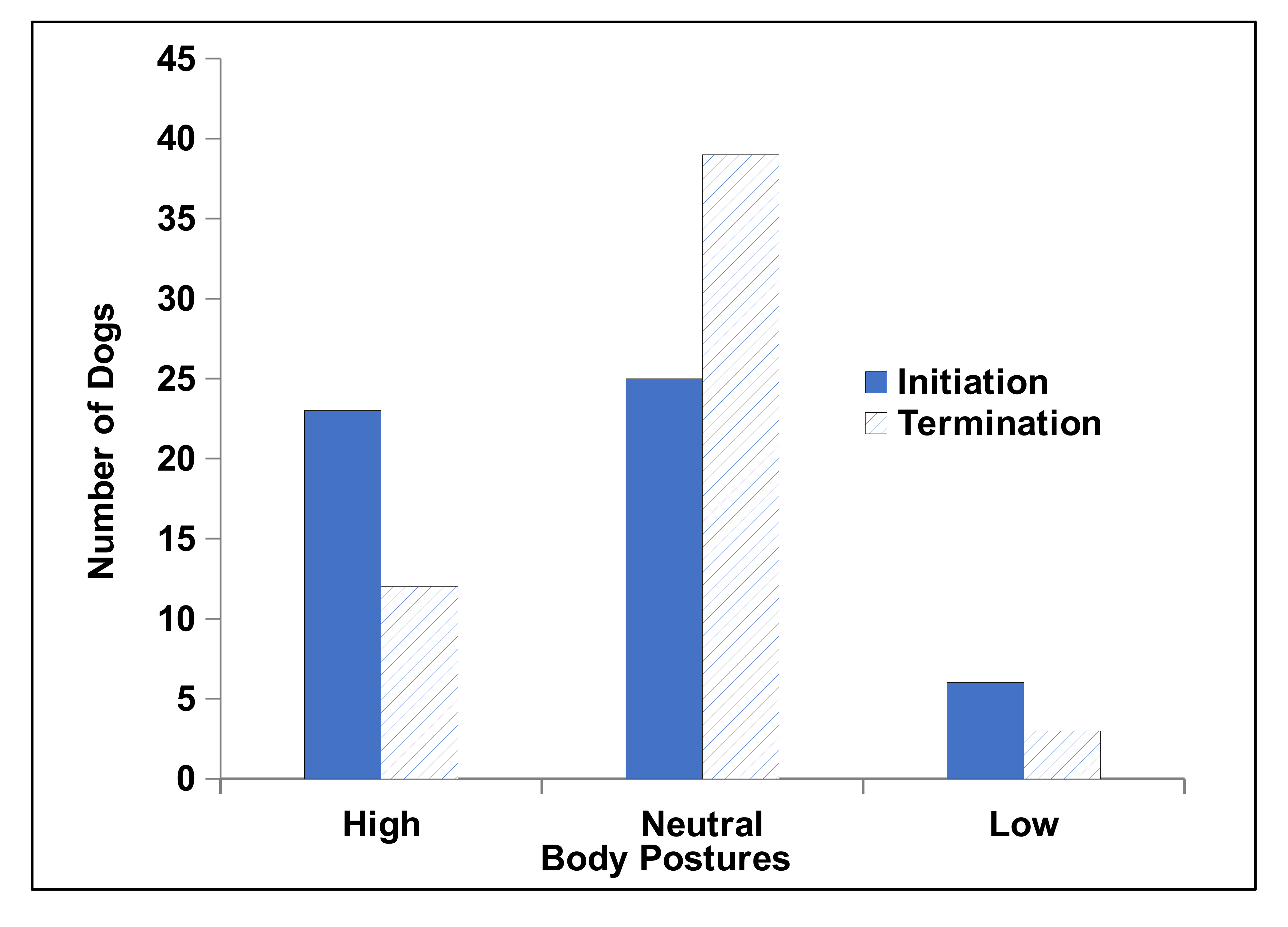
Figure 2. Number of dogs displaying “high”, “neutral”, and “low” body postures at the initiation of a greeting compared to body postures of the same dogs at the termination of the greeting. McNemar’s test: X22 = 10.563, N = 54, P = 0.001.
Olfactory Inspections
There was a total of 65 olfactory inspections across all dogs and dyads. Of the 16 dogs who shifted body postures when greeting initiation and termination postures were compared (see above), 87.5% had engaged in olfactory inspections during the same greeting (Binomial test: N = 16, P = 0.002).
The dogs initiating olfactory inspections in single-dog greetings varied among three groups: initiator, recipient, and both initiator and recipient (Chi-square test of goodness-of-fit: X22 =7.182; P = 0.028). Initiators engaged in a higher frequency of olfactory inspections (Binomial test: N = 13; P = 0.021) than expected by equal chance probability across groups. Recipient and both initiator and recipient groups engaged in olfactory inspections at chance levels (Binomial test for recipient: N = 3; P = 0.068; Binomial test for both initiator and recipient simultaneously: N = 6; P = 0.655).
In dual-dog greetings (N = 8), both initiators engaged in olfactory inspections during greetings in all cases.
For single-dog greetings, first olfactory inspections did not differ by body region, but results approached significance (Chi-square test of goodness-of-fit: X22 Χ = 5.545; P = 0.062). First olfactory inspections to the anogenital region, face/head, or inguinal regions occurred in 54.5%, 31.9%, and 13.6% of greetings, respectively. For dual-dog greetings, 66.7% of first olfactory inspections were to the face, but the small occurrence of dual-dog initiations (N = 6) precluded statistical analysis by body region.
Discussion
The current study offers new insights on greeting behavior in dogs that complements and adds to the comparative work on greeting behavior from both evolutionary and applied frameworks. I predicted that dogs would initiate greetings to maximize benefits and minimize costs to the initiator, which was the case. Single-dog greetings (where one dog initiated a greeting) were more common than dual-dog greetings (where both dogs initiated a greeting), and the initiator was often bigger, had body posture equal to or higher than the recipient, and was more likely to engage in olfactory inspections than the recipient. During a greeting, dogs gain olfactory information about the other dog (Howse 2018), and it’s safer to do so when the initiator is bigger and at least equal in status and confidence, as measured by body postures (Derix et al. 1993; van der Borg et al. 2015), to the recipient dog. In all cases of dual-dog greetings, both dogs initiated and engaged in olfactory inspections, further emphasizing the importance of the information-gathering component associated with greetings.
When off leash, and left to their own device, dog greet times were relatively short: 6.042 ± 0.801 s. Dogs investigated one another briefly and then departed, or infrequently, play ensued. A recent study on encounters between pairs of unfamiliar dogs at a dog park in Germany (Iotchev et al. 2019) reported that pairs spent 17.3% (~51.9 s) of a 5-min interval in proximity to one another (measured as two dog body lengths). In the Iotchev et al. study, dogs were pre-assigned to dyads and released into an unfamiliar fenced area (200 m2) rather than allowed to make a free choice about which dog to approach and greet. Additionally, the authors were not focused on initial meet and greet times, but instead reported time spent in proximity near the other dog during a pre-determined time interval. Results may vary according to testing conditions (observational vs. experimental), data collection methodology, and the dogs’ living conditions—i.e., pet vs. kenneled. For example, kenneled beagles spent more time in proximity to an unfamiliar conspecific than a familiar one during a 3-min interval (Pullen et al. 2013).
Length of greeting could also vary according to the purpose of the greeting—i.e., gathering information about unfamiliar, unrelated individuals vs. promoting cooperation and social bonds necessary for survival, as seen in African wild dogs greeting ceremonies (Smith et al. 2011). In captive African wild dogs, for example, greetings lasted up to 226 s during times when the alpha female was pregnant (Rütten and Fleissner 2004). Notwithstanding the variables that could affect greeting length, the results in this study suggest that dogs often engage in short greetings lasting only a few seconds.
After the initial short greeting, one or both of the dogs moved away without owner interference (with the exception of those few greetings that led to play). This finding is of applied importance as such: Owners walking their dogs on leash in urban or suburban areas may run into a friend on their walk and stop to talk. The friend also may have a dog on a leash. Now the two people are standing and talking and the leashed dogs are forced into close proximity for what may be an extended period that is beyond the comfort level of one or both of the dogs (often beyond the reported mean greet time in this study), and neither dog has the option to move away because their movement is restricted by a short leash. This may lead to reactivity such as barking or lunging, which then causes the owners to separate the dogs, inadvertently rewarding the aggressive display. This scenario is one that is all too familiar to behavioral clinicians working with dogs who are exhibiting problem behaviors (Overall 2013). Having ethological data on average greet times collected under semi-natural conditions (off leash, not hindered by the owner, dogs could physically move away, in an environment the dogs would normally meet, etc.), such as in the current study, is beneficial for applied animal behaviorists working with owners and dogs in the community and designing behavioral modification plans to address dog-to-dog aggression and reactivity.
In the current study, greetings rarely led to play (13.8% of the time), suggesting that dogs greet for reasons other than to engage in play. However, I never observed a greeting leading into aggression. Aggression is also reported as infrequent in other dog park studies (Shyan et al. 2003; Howse et al. 2018). One explanation is that if greetings between dogs function, in part, as a conflict management strategy, then aggression associated with greetings should be uncommon, as observed in the current study. A decline in the number of dogs whose body postures indicated arousal and fear at the end of the greeting compared to body postures at the beginning of the greeting lend support for this hypothesis (see discussion on body postures below). A second explanation is that dogs at the dog park are likely a self-selected group because dogs who are more likely to show aggression towards other dogs may be banished from the park (human peer pressure) or owners may be less willing to bring them to the dog park in the first place, as suggested by Bradshaw and Lea (1992) who also reported a rarity of aggressive behaviors in dogs meeting in open areas in the U.K. The explanations are not mutually exclusive, and both may contribute to the lack of aggression observed between dogs in the current study.
Do greetings help to reduce arousal and help dogs to return to a more “neutral” state? When body postures for the same dogs where compared at the initiation of the greeting and at the termination, those dogs who shifted postures moved from a state of arousal (high posture) or fear (low posture) at the beginning of the greeting to a neutral posture at the conclusion of the greeting. Interesting enough, I never observed any incidents of two dogs with low postures greeting and only one incident of a dog with a lower body posture initiating a greeting. Motivation to greet another dog is likely to be low if the individual is uncertain and/or fearful.
In both baboons and hyenas, greetings can help to reduce agonistic motivation (fear or aggression) and uncertainty between individuals and increase social tolerance (Colmenares 1990; Colmenares et al. 2000), and greetings can have an appeasing effect in some primates (Dunbar and Dunbar 1975; Hausfater and Takacs 1987). Greetings may, in part, be operating similarly in dogs. In a study examining temperament in dogs, De Meester et al. (2011) found that dogs with high body postures, in confrontation with another dog, were more likely to show aggression than dogs with low body postures, although dogs with low postures will use aggression too but to a lesser extent. The change in body postures for the same dogs between the beginning and end of greetings suggests that through greetings, dogs acquire information that reduces arousal and fear for some dogs. This further highlights the role that greeting behaviors may play in conflict management.
Research into how dog greetings affect arousal, fear, and aggression in different populations (dog-to-dog greetings vs. dog-to-human greetings, leashed vs. unleashed dogs, etc.) as well as identifying important variables related to greeting—e.g., when do greetings lead to aggression? —would be extremely useful information for applied companion animal practitioners. This study represents a first step in that direction.
Dogs in reciprocated greetings, where both dogs engaged in olfactory inspections, were more evenly matched weight wise compared with dogs involved in unreciprocated greetings. This suggests that in dyads where dogs are more evenly matched, both individuals may be interested in assessing the potential competition more often than in dyads where obvious discrepancies in size may make direct inspection less pressing. A bigger dog generally has a competitive size advantage over a smaller dog. As such, dogs in dyads with large discrepancies in size between greeters can make investigating vulnerable areas of a much larger dog potentially risky for the smaller individual. Although there was no aggression observed between dogs greeting in the current study, there are reported incidents of smaller dogs being attacked and killed at dog parks (ABC 10 News 2016). Additionally, the mean weight of dogs in this study was 25.670 ± 1.303 kg. Therefore, the results reported here are applicable to greetings involving primarily medium-sized or larger dogs.
The location of first olfactory inspection did not differ by body region, although there was a non-significant trend for dogs to investigate the anogenital region most often and the inguinal region least often. These results contrast with some previous studies, and discrepancies in findings may be related to differences in study design and data collection. For example, dogs meeting in two open areas in the U.K. directed olfactory inspections to the head and anal regions (Bradshaw and Lea 1992). However, Bradshaw and Lea included data from both leashed and unleashed dogs, whereas all dogs in the current study were off leash. In a different study, dogs entering a dog park in St. John’s Newfoundland were more likely to receive (rather than give) “snout-muzzle” contact. When the entering dog did make snout-muzzle contact, it was to the head region of the other dog(s) most frequently (Howse et al. 2018). However, Howse et al. used data collected within the first 400 s after the dog entered the park, whereas the current study did not incorporate data on dogs newly arriving to the park.
The current study provides fundamental information on dyadic greeting behavior between dogs visiting a dog park, but the findings reported here don’t necessarily generalize to all dog parks or public settings. Factors such as convenience sampling and small sample size (e.g., data from one park), as well as differences in the physical environment (e.g., park size) and social makeup of subjects (e.g., ages, sizes of subjects) among parks may result in findings that differ across locations. Despite these potential limitations, the current study offers new and valuable information that describes and enhances our understanding of greeting behavior between dogs in a dog-park environment.
Conclusion
This study found that single-dog greetings were more common than dual-dog greetings and that dogs initiated greetings to maximize gains and minimize costs associated with the behavior. This most often was the case as bigger dogs, and dogs with body postures equal to or higher than to the recipient were more likely to initiate greetings and to engage in olfactory inspections in single dog greetings.
Additionally, reciprocated greetings occurred more often when dogs were more evenly matched in terms of size as predicted.
Greetings allow dogs to gather information about conspecifics in ways that may decrease the risk of aggression. For example, body posture changes associated with greetings, from either a high or low posture at the initiation of a greeting to a neutral posture at the termination of a greeting, suggest that greetings play a role in conflict management by reducing arousal and/or fear that may lead to aggression in some dogs. This is the first study to demonstrate that greetings may function, in part, to reduce negative affect in some dogs, as measured by changes in body postures.
Greeting times were relatively short when dogs were allowed to greet and move away without interference of a leash. This is the first study, to my knowledge, that establishes a baseline for greeting times between dogs under semi-naturalistic conditions. This finding is relevant to animal behavior researchers, clinicians working in companion animal behavior, and dog owners alike.
Acknowledgements
I am grateful to all owners and dogs at Swift Run Dog Park who participated in the study and to an anonymous reviewer whose comments and suggestions helped to improve the quality of the manuscript.
Ethical Considerations
EU approval was not required in accordance with EU Direction 2010/63/EU for animal experiments. The study was fully observational involving dogs owned by private individuals.
References
ABC 10 News, 2016. Chihuhuhua dies in Ocean Beach dog park attack. Retrieved from http://www.10news.com/news/chihuahua-dies-in-ocean-beach-dog-park-attack.
Aureli, F. and Schaffner, C.M. 2007. Aggression and conflict management at fusion in spider monkeys. Biology Letters 3: 147-149.
Bauer, E.B. and Smuts, B.B. 2007. Cooperation and competition during dyadic play in domestic dogs, Canis familiaris. Animal Behavior 73: 489-499.
Beerda, B., Schilder, M.B.H., van Hoof, J.A.R.A.M., de Vries, H.W. and Mol, J.A. 1998. Behavioural, saliva cortisol and heart rate responses to different types of stimuli in dogs. Applied Animal Behaviour Science 58: 365-381.
Bekoff, M. 1984. Social play behavior. Bioscience 34: 228-233.
Bradshaw, J.W. and Lea, A.M. 1992. Dyadic interactions between domestic dogs. Anthrozoös 5: 245-253.
Colmenares, F. 1990. Greeting behavior in male baboons. I: communication, reciprocity and symmetry. Behaviour 113: 81-116.
Colmenares, F. 1991. Greeting, aggression, and coalitions between male baboons: demographic correlates. Primates 32: 453-463.
Colmenares, F., Hofer, H. and East, M.L. 2000. Greeting ceremonies in baboons and hyenas. In Natural Conflict Resolution, 94-96. eds. F. Aureli and F.B.M. de Waal. Berkley: University of California Press.
Deldalle, S. and Gaunet, F. 2014. Effects of 2 training methods on stress-related behaviors of the dog (Canis familiaris) and on the dog-owner relationship. Journal of Veterinary Behavior 9: 58-65.
De Marco, A., Sanna, A., Cozzolino, R. and Thierry, B. 2014. The function of greetings in male Tonkean macaques. American Journal of Primatology 76: 989-998.
De Meester, R.H., Pluijmakers, J. and Vermeire, S. 2011. The use of the socially acceptable behavior test in the study of temperament of dogs. Journal of Veterinary Behavior 6: 211-224.
Derix, R., van Hoof, de Vries, H. and Wensing, J. 1993. Male and female mating competition in wolves: female suppression vs. male intervention. Behaviour 127: 141-174.
Dias, P.A.D., Luna, E.R. and Espinosa, D.C. 2008. The functions of the “greeting ceremony” among male mantled howlers (Alouatta palliata) on Agaltepec Island, Mexico. American Journal of Primatology 70: 621-628.
Dunbar, R.I.M. and Dunbar, E.P. 1975. Social dynamics of gelada baboons. Contributions to Primatology 6: 1-157.
Fagen, R.M. 1981. Animal Play Behavior. New York: Oxford University Press.
Harrington, F. H. and Asa, C.S. 2003. Wolf communication. In Wolves: Behavior, Ecology, and Conservation, 66-103, eds. L.D. Mech and L. Boitani. Chicago: University of Chicago Press.
Hasegawa, M., Ohtani, N. and Mitsuaki, O. 2014. Dogs’ body language relevant to learning achievement. Animals 4: 45-58.
Hausfater, G. and Takacs, D. 1987. Structure and function of hindquarter presentations in yellow baboons (paio cynocephalus). Ethology 74: 297-319.
Hecht, J. and Horowitz, A. 2015. Introduction to dog behavior. In Animal Behavior for Shelter Veterinarians and Staff, 5-30. eds. E. Weiss, H. Mohan-Gibbons, and S. Zawistowski. Ames: John Wiley and Sons.
Horowitz, A. 2009. Attention to attention in domestic dog (Canis familiaris). Animal Cognition 12: 107-118.
Howse, M. 2018. Social behavior of domestic dogs (Canis familiaris) in a public off-leash dog park. Behavioural Processes 157: 691-701.
Iotchev, I.B., Egerer, A., Grafe, S., Adorjan, A. and Kubinyi, E. 2019. Encounters between pairs of unfamiliar dogs in a dog park. Biologia Futura 70: 156-165.
Kiddie, J.L. and Collins, L.M. 2014. Development and validation of a quality of life assessment tool for use in kenneled dogs (Canis familiaris). Applied Animal Behaviour Science 158: 57-68.
Kummer, H. 1968. Social Organization of Hamadryas Baboons. A Field Study. New York: Karger.
Kutsukake, N., Suetsugu, N. and Hasegawa, T. 2006. Pattern, distribution, and function of greeting behavior among black-and-white colobus. International Journal of Primatology 27: 1271-1291.
McDonald, J.H. 2008. Handbook of Biological Statistics. Baltimore: Sparky House Publishing.
McHugh, M. 2012. Interrater reliability: The kappa statistic. Biochemical Medicine 22: 276-282.
Nakagawa, S. 2004. A farewell to Bonferroni: the problems of low statistical power and publication bias. Behavioural Ecology 15: 1044–1045.
Overall, K. 2013. Manual of Clinical Behavioral Medicine for Dogs and Cats. St. Louis: Elsevier.
Pageat P. and Gaultier E. 2003. Current research in canine and feline pheromones. Veterinary Clinics of North America: Small Animal Practice 33: 187-211.
Perinetti, G. 2018. StatTips Part IV: Selection, interpretation and reporting of the intraclass correlation coefficient. South European Journal of Orthodontics and Dentofacial Research, 5: 3-5.
Petrak, I. 2018. Patterns of carnivores’ communication and potential significance for domestic dogs. Periodicum Biologorum 112: 127–132.
Preuschoft, S. and Van Schaik, C.P. 2000. Dominance and communication: conflict management in various social settings. In Natural Conflict Resolution, 77-105. eds. F. Aureli and F.B.M. de Waal. Berkley: University of California Press.
Pullen, A.C., Merrill, R.J.N. and Bradshaw, J.W.S. 2013. The effect of familiarity on behavior of kenneled dogs during interactions with conspecifics. Journal of Applied Animal Welfare Science 16: 64-76.
Řezáč, P., Viziová, P., Dobešová, M., Havlíček and Pospíšilová, D. 2011. Factors affecting dog-dog interactions on walks with their owners. Applied Animal Behaviour Science 134: 170-176.
Rütten, S. and Fleissner, G. 2004. On the function of the greeting ceremony in social canids—exemplified by African wild dogs Lycaon pictus. Canid News 7.3 (online). Retrieved from http://www.canids.org/canidnews/7/Greeting_ceremony_in_canids.pdf.
Schenkel, R. 1967. Submission: It’s features and function in the wolf and dog. American Zoologist 7: 319-329.
Shyan, M.R., Fortune, K.A. and King, C. 2003. “Bark parks”—a study on interdog aggression in a limited control environment. Journal of Applied Animal Welfare Science 6: 25-32.
Siniscalchi, M., d’Ingeo, S., Minunno, M. and Quaranta, A. 2018. Communication in dogs. Animals 8: 131, accessed December 02, 2019, doi:10.3390/ani8080131.
Smith J.E., Powning, K.S., Dawes, S.E., Estrada. J.R., Hopper, A.L., Piotrowski, S.L. and Holekamp, K.E. 2011. Greetings promote cooperation and reinforce social bonds among spotted hyaenas. Animal Behaviour 81: 401-415.
Smuts, B.B. 2002. Gestural communication in olive baboons and domestic dogs. In The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition, 301-306, eds. M. Bekoff, C. Allen, and G.M. Burghardt. Cambridge: MIT Press.
Smuts, B.B. and Watanabe, J. M. 1990. Social relationships and ritualized greetings in adult male baboons (Papio cynocephalus anubis). International Journal of Primatology 11: 147-172.
Trisko, R.K. and Smuts, B.B. 2015. Dominance relationships in a group of domestic dogs (Canis lupus familiaris). Behaviour 152: 677-704.
The Trust for Public Lands. 2018. The dog park rankings for the 100 largest U.S. cities, 2018. Retrieved from https://cloud-tpl.s3.amazonaws.com/images/landing-pages/ccpf/2018/City%20Park%20Facts_Dog%20Parks%202018.pdf.
van der Borg, J.A.M., Schilder, M.B.H., Vinke, C.M. and de Vries, H. 2015. Dominance in domestic dogs: A quantitative analysis of its behavioural measures. PLoS ONE 10: 1-18.
van Hooff, J.A.R.M. and Wensing, J.A.B. 1987. Dominance and its behavioral measures in a captive wolf pack. In Man and Wolf: Advances, Issues, and Problems in Captive Wolf Research, 219-232, ed. H. Frank. Dordrecht: Dr. W. Junk Publishers.
Wan, M., Bolger, N. and Champagne, F.A. 2012. Human perception of fear in dogs varies according to experience with dogs. PLoS ONE 7: e51775.